In this chapter, Roberts concludes her survey of human evolution and development with final remarks.
Chapter Summary:
In the concluding chapter of her book, Roberts surveys the history of understandings of our own development and evolution. She recalls previous misinterpretations of our origins and looks to the future of science to further explain the things that make us human. She returns to her focus on embryology to point to the shared history that is so obvious in our development and points out that many anatomical differences arise through tinkering with our already present set of genes during gestation which ultimately changes how our body will develop over our lifetime. Evolution is constrained by what already exists. This allows us to see our history in other species that led to our own development and to see how we have modified our body plans over the years yet maintain many similarities with other vertebrates and indeed all living creatures. We create the technological and social environment we inhabit, and this will inevitably lead to changes in ourselves. Things change rapidly as technology progresses and we will continue to change, just as our ancestors did in response to the use of stone tools. Humans do not have traits that are entirely unique to us, rather it is the combination of traits that leads to our own uniqueness. Every species is different, we do not hold a privileged position in the tree of life as the epitome of evolution. Our future is full of responsibilities and Roberts asks readers to accept this responsibility to ourselves and the entire planet and to fight for a sustainable future.
In this chapter, Roberts addresses the unique physiology of the human shoulder and hands and how they reflect our evolutionary past. She also talks about the things we share with other apes as well as what sets us apart and why these differences may have arisen.
Chapter Summary:
Human arms and hands are much more mobile than those of most other mammals. Our shoulder anatomy is owed in part to our climbing ancestors. We have free-floating scapulae and very mobile arms. Shoulders are a very mobile and as a result of this mobility, they dislocate more easily than more stable joints. Human forearms are also very mobile. They have the ability to pronate which allows rotation around the arm-hand axis. Other apes have more upward facing shoulders because of their use for climbing. Roberts discusses possible reasons that our shoulders have rotated downwards. Perhaps they conferred an advantage for long distance running. Some suggest that the change in shoulders may be because of advantages for throwing. This seems to have been very important because much of our anatomy seems well suited for throwing. There is little fossil evidence of throwing before the invention of tools specifically designed as projectile weapons such as throwing spears. There is conclusive evidence that throwing was used for hunting by 300,000 years ago.
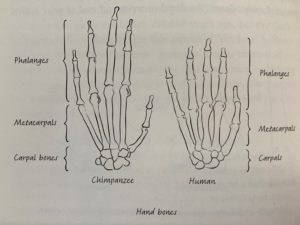
Skeletal differences between human and chimpanzee hands show that our hands serve different functions despite their seeming similarity (Roberts 324).
Next, Roberts discusses the ability of human and ape wrists to be in both supinated and pronated positions allowing us to use our hands for even more things. Our hand bones are different than those of chimps in very significant ways. They have short thumbs and long phalanges compared to our more modest phalanges and large, strong thumbs. Chimpanzees use their fingers to grip sticks while humans use their thumbs to allow for greater control and precision. Humans use their hands with a combined precision and strength grip that allows us greater dexterity with our hands.
Ardi’s fossils are useful for determining when hands diverged between humans and other apes. Longer fingers are associated with grasping tree branches to hang from. Evidence suggests that metacarpals have elongated in apes while they have shortened in humans since our last common ancestor. Curved finger bones in Lucy and her species suggest that they spent some time in trees, as well as walking bipedally. Her spine suggests that this amount of time was not great because it is very flexible and much better adapted to walking than climbing where she may risk injury. By Homo habilis hands were more similar to modern humans, though more robust than modern humans.
We also have different musculature in our hands than other primates. There are three muscles that are unique to the human hand and all of them flex the thumb in different ways, underlying our unique use of our thumb for precision and strength gripping. Some suggest that these muscles are a result of toolmaking. This is true for the robust thumbs we see in earlier hominins too, which may have been good for the high impact methods used to create tools. Another theory suggests that our robust thumbs are a result of pressures experienced when using stone tools instead of when they were being made.
In this chapter Roberts discusses the evolution of human bipedalism as well as its modern form and implications of physiological plasticity in modern humans as a potential feature that can be selected for.
Chapter Summary:
In this chapter, Roberts discusses the development of bipedalism in humans. This is one of the few things that sets us apart from our primate relatives. A few other tetrapods share this strategy with us but overall it is a pretty strange adaptation. Many parts of the environment interact with genes to shape our physiology. These influences include diet and stress. One of the earliest signs for the transition to bipedalism is movement of the foramen magnum. This allows the head to sit in a more upright position. Evidence of this may exist in as early specimens as Toumai which lived 6 to 7 million years ago. Later fossils show more conclusive evidence of bipedalism. Judging from the hips of her species, it seems they had a human-like form of locomotion. Attachments for hip abductor muscles are visible in the pelvic processes in us and earlier bipeds. These muscle attachments allow for our easy gait and keep our opposite foot from dragging on the floor when we take a step. Evidence from Lucy’s spine suggests that she had a spine that curved like ours and allowed her to balance her weight on one leg as she took steps.
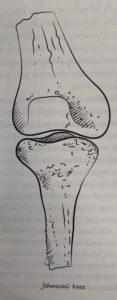
The anatomy of a Johanson’s knee is thought to allow for better balance during bipedal locomotion (Roberts 286).
There is some evidence that Lucy had a Johanson’s knee which is a knee that curves inward so that the foot that is on the floor is closer to the center of gravity and allows balance while standing on one foot. A sloping femur and angled knee help to give us the balance we enjoy.
Human ankles are specially shaped to support our weight and are very good at moving along the axis of movement that they need to support efficient locomotion. Fossilized footprints show that hominins were walking upright 3.7 million years ago. Distribution of weight from these footprints corroborates the previous theory that Australopithecus aferensis (Lucy’s species) was able to walk with a fairly efficient gait. We are not as physiologically limited for tree climbing as we may think. If humans climb trees often and from a young age, the soft tissues may form in a way that allows much greater flexibility in the ankle joint and allows us to both walk efficiently and to climb trees similarly to how apes, especially chimpanzees can. We need to be careful to not draw too harsh of conclusions from bones because soft tissues also play a large role in determining functionality of structures.
Roberts cautions against falling into thinking that our mode of locomotion developed linearly through time. She suggests it probably had some other use first then was reapplied to the ground, in contrast to the common narrative of knucklewalking apes beginning to become more and more upright. Roberts suggests that apes that walked in the trees on two legs sometimes were the first step towards upright walking on the ground. Perhaps we are the ones with an old trait and gorillas and chimpanzees have come up with the new mode of transportation in knucklewalking.
Not all of our leg physiology can be explained by walking. In fact, in H. erectus specimens, there is significant evidence suggesting that many of the adaptations favor efficient running. Perhaps this is why our legs are so long, to allow for efficient endurance running. Human adaptations to running are an excellent example of how the environment can shape physiology before genes change. Roberts describes the Baldwin Effect which suggests that variation can arise during the lifetime and these changes can be passed on to offspring in the form of epigenetics. This variation is selected for genetically, but more indirectly, these pressures select for genes that can lead to the desired phenotype. In this way it is important to have genes that are activated to be affected by evolution. Latent potential to be able to pick up new forms of locomotion may be an important factor selected for by evolution even more than direct physiological adaptations.
In this chapter, Roberts discusses the emergence of limbs in human embryos as well as how they probably evolved as fish emerged from the water.
Chapter Summary:
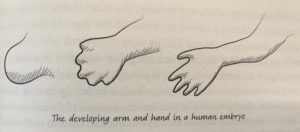
Roberts begins her discussion of limbs by describing their embryonic origins. Starting as limb buds early in development, they will develop into our arms and legs. By week seven, fingers and toes begin to form.
The development of fingers in a human embryo through apoptosis during early development (Roberts 265).
By week six limbs contain cartilage models of bones which will form as the fetus develops. Bones continue to ossify during growth. Growth plates in different bones fuse at different developmental stages in life. For osteoarcheologists, this gives tools to make a good guess at age when looking at the remains of a juvenile.
Muscles migrate to their attachment points during embryonic development. Somites originating from near the vertebrae attach and fuse together to make the muscles adults have. Patterns of innervation show the migration patterns of muscles from the spinal cord.
Fins and limbs share an evolutionary history. There are many similarities between arms, fins and wings of animals. Homologous structures show this shared past. Roberts discusses the controversial anatomist Richard Owen and his own description of these similar structures, though he was a firm believer in the scala naturae, these observations were useful in seeing patterns in our history and call back to the Hox genes that we rely on for body planning. One mutation could have caused the addition of fingers or toes in our ancestors, a much more credible transition than developing an entire new extremity from scratch.
Next, Roberts discusses the emergence of limbs in a fishlike ancestor, the Acanthostega which shared many characteristics with tetrapods as well as fish. This creature, however, showed a strange number of fingers, eight on each forelimb and probably as many on its hind legs. Contrary to logic though, it seems these limbs were not useful for bearing weight. It is thought that these creatures waded in shallow waters and were able to propel themselves out of the water briefly to capture prey then they waded back into the water where they spent most of their time. Another fossil, Tiktaalik, which similarly was a transitionary animal between fish and land animals, shows evidence that pelvic fins, which were traditionally thought to be weaker than pectoral fins (which would become the forelimbs of modern tetrapods), had begun to become stronger, as reflected with the comparative robustness of the pelvic bones of the Tiktaalik. This strength shows that perhaps hind fins in transitionary animals were already making the shift to hindlimb driven locomotion which is what we see in the stronger hind limbs of most tetrapods now.
Roberts discusses the reproductive systems of humans and our closest relatives to explore the somewhat unique features and challenges of human reproduction.
Chapter Summary:
This chapter is a survey of the human reproductive system. It begins with a description of the goal of the reproductive system, to unite a sperm and egg and to form a new human. Transferring from water to land was a challenge for reproduction. Most fish release their eggs then another fish releases its sperm above the nest to fertilize them. Land animals have a harder time. One way that animals have overcome this is by returning to the water to lay their eggs like frogs and other amphibians. Reptiles began to engage in the first coital transfer of gametes. A male and female reptile press their cloaca (opening for the large intestine, bladder and reproductive tracts) together and the sperm is transferred from the male into the female. The fertilized egg is then deposited by the female after developing a little. Birds and mammals are descended from this line of reproductive strategies. Most birds use a similar strategy to reptiles to transfer sperm, but a few reptiles and birds have erectile penises. Mammals have external penises which they use to introduce their sperm into their mate.
In embryos, both female and male genitals follow a similar developmental pattern. They start the same and eventually the X or Y chromosomes are activated, and the genitals will differentiate into those of the sex of the fetus. In the end, the reproductive organs of males and females look quite different, but they are made up of largely the same tissues. The evolutionary history of the testes involves a duplication of the kidneys called the mesonephrons. In humans, these structures become the Wolffian ducts and descend to the testes and become the sperm ducts of the testes and the vas deferens, connecting the reproductive tract with the urinary tract. In women, these structures do not play a part in reproductive anatomy. In females, the Mullerian ducts are dominant and become the oviducts of the female reproductive system while the Wolffian ducts disappear. One strange feature of the human female reproductive system is the presence of a single, unforked uterus which allows for us to sustain a pregnancy and have single births.
Next, Roberts discusses the migration of gonads in both females and males. In males, the gonads migrate a long distance to rest in the scrotum. In females they descend from their original position in the back of the abdominal cavity to where they will rest above the uterus. Roberts then discusses the vulnerable position of the testicles in human males. She explains that it is thought that sperm production with the least error in DNA production and with the greatest efficiency mandates lower temperatures. Finally, Roberts discusses the size of human testicles and how they compare to our closest relatives, chimpanzees and gorillas. Gorillas, though they have the largest bodies, have the smallest testicles. And chimpanzees, though they have the smallest body size possess the largest testicles. This has to do with sexual competition. In highly promiscuous species like chimps, females may have the sperm of more than one male in their reproductive tracts so having more sperm per ejaculation and higher quality sperm gives a male a higher chance of fertilizing an egg. Gorilla males do not compete with other males directly. If they have a harem of females, they have exclusive mating rights to the group. In this way they do not need as much sperm to ensure fertilization. Humans, in the middle of these extremes, have a mix of promiscuity and monogamy which is reflected in our testicles.
Roberts discusses the anatomy of the human penis and clitoris. The penises of mammals are widely varied, and the clitorises of mammals are similarly varied. The mechanisms that result in an erection in males are very similar to those that are involved with the female erection. Erections are the result of an increase in the production of cGMP in the area which causes arteries to swell and increase pressure in erectile tissues. In erectile dysfunction, the enzyme that breaks down cGMP in the genitals breaks down cGMP before it can take full effect. The drugs that correct this dysfunction in males may also be a promising lead towards helping to alleviate similar dysfunction in females.
Oxytocin is an important chemical in pair bonding as well as bonding between parents and children and is also important for biological functions including orgasm and milk letdown. The female orgasm is still not understood well, this lack of research stemming largely from cultural misrepresentations and societal ideals. In females, orgasm seems to be triggered by stimulation of the vagina and clitoris, the G-spot, if it exists, may also be important. Humans are reproductively active at all times of year. Because of our joint mobility, we can have sex in a greater variety of positions than other mammals. Sexual behavior is highly social. In many other primate species, sexual acts with members of the same or the opposite sex occur frequently and seem to serve social purposes such as reducing tension.
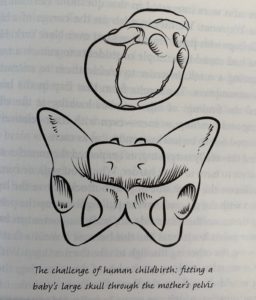
The tight fit between the skull of a baby and the pelvis of a modern human mother (Roberts 252).
Finally, Roberts discusses difficulty birthing for humans. She explains the obstetric dilemma, the opposing pressures for cranial development and pelvic width restrictions because of walking efficiency. She then describes the faults of this theory. Metabolically, it is not more expensive for people with wider hips to walk. And furthermore, humans have very long gestational periods despite having relatively helpless youth. She proposes the energetic crisis hypothesis. This states that the mother cannot increase her metabolism quickly enough to keep up with the growth of the child, so birth occurs. Even though the birthed baby requires more energy (in the form of milk) than the gestating baby, the rate of increase slows after birth, perhaps allowing the mother to continue to keep pace with the growing young. Perhaps, she suggests, the truth is a mix of different reasons. Perhaps our cooperative birthing system has lessened the evolutionary pressure to widen hips and allowed for us to continue surviving difficult births. Whatever the truth is, surgical intervention during birthing allows for us to continue to avoid strong evolutionary pressures for wider hips and we may see these consequences exacerbated in the future because of it.
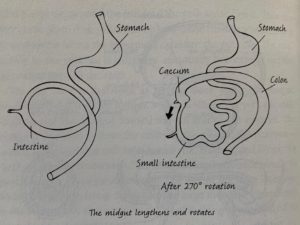
Roberts discusses the development of the gut during development and the evolutionary roots of our modern gut physiology as well as whether it is unique or standard for a primate of our size and diet.
Chapter Summary:
The development of the human gut in embryos outside of the body creates our adult pattern of gut organization (Roberts 212).
Guts all originate in the same way, as a simple tube of endoderm. Human embryos have a yolk sack, a reminder of our close relation to other animals who lay eggs. The yolk sac in humans is not useful as embryos and fetuses get their nutrients from their mother and it degenerates by the time of birth. The development of the gut starts simply but the tube convolutes and specializes depending on region to create the entire gut from the mouth to the anus. The gut grows so quickly that the abdomen does not have time to keep up and the gut must herniate into the umbilical cord. This resolves as the fetus develops and the gut usually returns to the inside of the abdomen. In fact, this herniation creates the organization we have in our adult gut.
Roberts describes her own experience of having an exploratory procedure of her small intestine. Her reflections of the experience of having a small “pillcam” photograph her intestine describe how modern imaging techniques allow us to visualize areas of the body we would not be able to otherwise.
The human gut is rather standard as far as primates’ intestines go. Ours guts are unspecialized; they are capable of digesting a wide variety of food sources. Human guts are small for our body size. This may be because the gut is very energetically expensive. As the brain expanded, our gut may have contracted. This is thought to be a product of the ability of our brain to find high quality food which mandates less surface area to extract sufficient nutrients. Theories propose that the expansion of our brains and the beginning of food processing, the gut became shorter. Roberts denies the importance of these differences, suggesting that we really do have very standard primate digestive systems and that in the past we were too intent on identifying parts of ourselves that are unique.
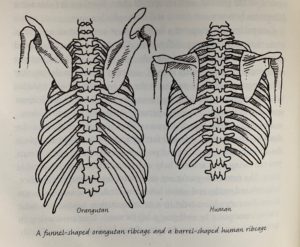
In this chapter, the shape of the chest and its contents are discussed from an evolutionary point of view, drawing on evidence from our ancestors and relatives to show how the anatomy of the chest may have arisen.
Chapter Summary:
This chapter begins with a description of the muscles and bones of the chest. The function of the intercostal muscles is to resist changing shape when we breathe in. The diaphragm and intercostal muscles oppose to create a negative pressure to suck air into the lungs so that we may breathe.
The shape of the ribcage is distinct from apes which swing from trees. Our chest shape may be an adaptation to bipedalism (Roberts 186).
Humans and closely related apes have a very special shape to our thoracic cavity. Our shoulders are laterally displaced, and our chests are flattened on the anterior-posterior axis. This allows us to swing our arms for an easier and more energy efficient gait. The attachment of abdominal rectus muscles allows us to move our trunk with considerable strength. This also affects the shape of the ribcage. Because of the advantages associated with barrel chested anatomy for walking bipedally, we may be able to look back in the past to see where obligate bipedalism became the only way to locomote for our ancestors.
The history of our chest shape is complicated. Australopithecus sediba specimens show more shrugged shoulders and apelike posture than Australopithecus afarensis which was an earlier specimen. Roberts cautions against thinking of evolution as a linear process that inevitably resulted in our current morphology. Instead, these are more hints about the ways our ancestors lived than about how or why we developed the chest we did. It may be that we have a chest more similar to our ancestors while other primate species are the ones who have changed.
Our chest shape is distinct from that of Neanderthals which allows us to see some of the distinctions between us and Neanderthals. Perhaps the larger thoracic volume of Neanderthals could show that they had larger lungs than modern humans do, possibly as an adaptation to altitude. Neanderthals had a more traditional robust body shape than Homo sapiens. In fact, it was H. sapiens that changed from the original morphology. Neanderthals were well suited for cold climates, but we are not sure that their morphology arose in response to cold climates.
Our heart is much the same as that of other mammals. We have a single loop circulation while we are fetuses but once we begin to breathe, we need a dual loop system which is why our hearts are split into the halves they are. Blood can flow from one side of the heart to the other in infants because blood mixing is unimportant, they do not absorb oxygen through their lungs. In adults this would lead to a very inefficient system of blood oxygenation. When this hole fails to close, it is usually corrected by a simple surgery.
Lungs develop much later than hearts during gestation because they are not needed until after gestation is complete. Problems with lung development arise when babies are born prematurely. This happens for two reasons. The alveoli are not yet mature and capable of gas exchange and surfactant in the lungs that allows for reduced surface tension is not produced yet. Without these two things ready, premature babies inevitably face problems with respiration.
The origin of a lung for breathing may lie in the swimming bladder in fish that allows them to float well. Lungfish are thought to be a good example of what the first air breathing animals looked like.
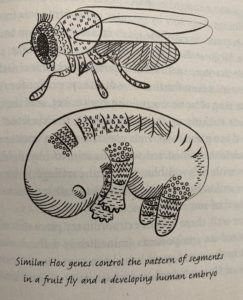
This chapter looks to genetic evidence from our distant relatives to show the conservation of Hox genes across all segmented creatures and discusses the human spine as a product of our evolution.
Chapter Summary:
Hox genes are shared between diverse species and control body organization and segmentation (Roberts 155).
Roberts opens this chapter by discussing the breakthroughs of genetics that fruit fly models were vital for by detailing the discovery of Hox genes using fruit fly models. Hox genes have an extremely ancient history. They exist in all segmented organisms. Even in distantly related species, these genes are shared and important for the layout of the body. She discusses that even snakes, which seem very different than humans and have many more vertebrae than us, share a common pattern of Hox genes.
The embryonic development of the spine is discussed next. The organization that takes place during embryonic development is very important for later function. In the embryo muscles migrate to more distal regions; their origin is visible in the nerves that connect to them. Roberts mentions the danger of spina bifida if there is insufficient folic acid while the spinal cord develops.
The anatomy of the spinal cord is complex. In humans, vertebrae have multiple important functions: weight bearing, protection for the spinal nerves and leveraging processes to allow for functional flexibility. Though individual vertebrae in humans are largely similar, there are two in particular in humans that have unique properties that allow us to support our large heads and move our necks so flexibly. The top two cervical vertebrae have a unique morphology that increases the mobility of the head along several axises of motion.
The number of each type of vertebrae (cervical, thoracic, lumbar, and sacrococcygeal) is different in different vertebrates. There are almost always seven cervical vertebrae in primates, though the number of caudal vertebrae is highly variable by species. Intervertebral disks are very unique as far as joints go. They have unique properties that allow them to absorb shock as well as have a wide range of motion. Though vertebrae are very good at being flexible, in humans they are vulnerable to injury, especially as we age. Pain perception is variable by region of the body. Topically, we are very aware of where our pain originates but internally, we have a much more difficult time locating the source of pain. This has to do with the organization of nerves off of the spinal cord.
Though flexibility in vertebrae has drawbacks, it is vital for our bipedal locomotion. The shape of our spine is a very important adaptation for upright walking and in the fossil, record serves as a very useful indicator of bipedal locomotion. Neanderthals, in contrast had straight spines. Evidence suggests that they were obligate bipeds, so it may seem counterintuitive that they developed stiff backs which detract from their ability to walk upright. Roberts suggests that this could be because they were able to benefit from the lower spine stability that comes with a stockier, less flexible spine. Maybe they carried things that were heavier or engaged in more taxing activities than our ancestors.
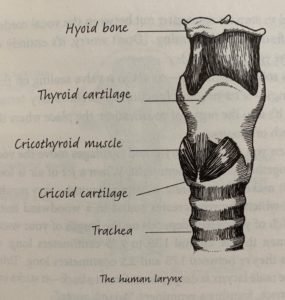
This chapter discusses the evolution of speech and all of the structures that develop from the branchial arches, which would be gills in other animals. Roberts takes an evolutionary approach to how these structures formed and how it is reflected in our evolutionary relatives.
Chapter Summary:
Roberts opens the chapter by discussing the physiology of modern human speech structures and how they compare to that of a Neanderthal skeleton found in Kebara. She discusses the complex structure of the larynx and the role of the hyoid bone in speech production. The larynx of humans is not unique, though. Many mammals have similar larynges. It is the structures above the hyoid that produce the specific sounds of speech. She discusses how the hypoglossal nerve tract was thought to give us an approximation of the mobility of the tongue, but later investigation found that this theory did not provide real hints about the ability to speak. It seems that ultimately, we will never know when humans developed the ability to speak, but there is evidence that Neanderthals were not as capable of making distinguishable vowels because of the length of their palate.
The anatomy of the modern human larynx and surrounding tissues (Roberts 119)
Roberts then discusses the myths surrounding the human larynx. It is not, in fact, perfect. She suggests that it is possible that the location of our larynx, which allows for us to speak clearly, could be a lucky accident of evolution in response to bipedalism. She suggests that if it had not descended, there would not be enough physical space in our throats for the larynx. Roberts claims that the physiological adaptations that allow for speech may not be indicative of the ability to speak at all, instead she proposes we look at the brain for hints about our use of speech.
Next, Roberts analyzes the lowered position of the adult male larynx in humans and why it may have evolved. She compares this to the voice box of male red deer, which is also much lower than the females of their species. Perhaps, like in red deer and other mammals, lower voices in human males are a product of sexual selection and sexual competition.
Roberts delves into the topic of the origin of the larynx. Surprisingly enough, it shares an evolutionary history with the gills of fish. In human embryos, structures resembling gills form and later become the larynx. She discusses the strange anatomy of the laryngeal nerve and points to its strange anatomical features as evidence of its evolution. The laryngeal nerve curves around the aorta and does a U-turn to come back up to innervate the muscles of the larynx. This is evidence of the migration of the heart in embryos and this reflects our evolutionary history.
Next there is a discussion of the ossicles of the ear and how the evolved from the duplication of the jaw hinge. Eventually these migrated and became a part of the auditory system. Roberts also adds a discussion of the evolution of human facial muscles that are useful for expressing emotions. She then describes the history of our earhole as a product of the first branchial cleft. The Eustachian tubes allow us to equalize the pressure of our inner ears and the interconnectedness of the anatomy of the head is clear in how seemingly unrelated activities such as yawning, or swallowing can open the Eustachian tubes. Our evolutionary history is better understood in light of its shared features with all sorts of animals.
The development of the physiology of the head is discussed in this chapter. Roberts uses evidence from embryonic development and our relatives to describe a narrative of how our head developed to look the way it does and the unique physiology of its sense organs.
Chapter Summary:
Roberts begins her discussion with a description of the neural crest, how it forms and how it will later be vital to the formation of the skull and many other important structures. She discusses the investigation of the evolution of neural crest cell genetics by looking at lancets and jawless fish, which are a closer relative to lancets but do possess neural crest cells during development. The difference, she explains, lies in large scale duplications of Hox genes which can then evolve to fulfill new functions, as is the case in vertebrates which have several copies of a particular Hox gene which is vital to the activation of neural crest cells in developing vertebrate embryos. She also talks about the role that neural crest cells can play after migrating, including having a vital role in the formation of the skull, an idea that was widely disbelieved until better methodology developed to prove that this was indeed the case.
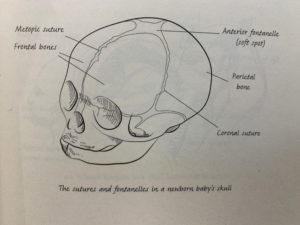
The human infant skull is very flexible to allow for an easier birth and brain expansion during development (Roberts 85).
Although the adult skulls of humans may look very different than those of other animals, in our infancy we have many similar structures that make our relationship easily identifiable. The malleability of the infant skull is very useful because it allows for expansion of the skull with growth of the brain. Culturally, this malleability has been taken advantage of culturally in many societies to form the skull by manipulation to socially desirable shapes. Cultural manipulation of the skull is not associated with impeding the brain from developing, but early closure of sutures can cause deformation of the brain.
Roberts starts a discussion of the general anatomy of skulls with the differences between male and female skulls, males usually having more robust features and women usually having a more gracile skull. She then discusses how remains can be aged using the sequence of teeth maturity or more roughly, suture closure in the skull and tooth wear. She then describes the general anatomy of the auditory system and how it transforms sound waves into nervous impulses. She discusses the olfactory system in mammals and the evolutionary degeneration of primates’ olfactory system compared to other mammals who are much more sensitive to smell. One theory is that this disinvestment came with a stronger reliance on vision. She discusses the seemingly impossible development of the eye. She describes that indeed the beginning of this complex structure probably was more like a photosensitive cone that developed later into more controlled and complex structures that we now use in our eyes. She proposes how the ability to sense light may have been advantageous in free swimming animas even before it allowed sight as we experience it today. Eventually this led to the development of eyes as we know them. Even a structure as complicated as the eyes can be reasonably interpreted in light of our developmental and evolutionary history.
Next, she discusses the history of color vision in mammals. In jawed fish, there are four different cones, allowing them to see in color. However, in most mammals, we see that there are only two, not allowing color vision. Only primates have regained this ability through evolution. Roberts proposes that this history points to a nocturnal ancestor that did not need color vision to see in the dark, then points to our recent history of fruit eating or predator spotting as a reason we may have regained this ability. She also discusses the placement of primate eyes, why ours are located where they are and how this is helpful for stereoscopic vision. She also talks about the large sclera of humans, which allows us to see where each other are looking in contrast with other animals which show little or no sclera. This is related to our social nature and brain development. Our ability to infer what each other think is very important for our sociality. She also discusses the physiology of the eye and the logical shortcomings that evolution has filled in with elaborate processing mechanisms.