
Chapter Overview:
In this chapter Roberts discusses the evolution of human bipedalism as well as its modern form and implications of physiological plasticity in modern humans as a potential feature that can be selected for.
Chapter Summary:
In this chapter, Roberts discusses the development of bipedalism in humans. This is one of the few things that sets us apart from our primate relatives. A few other tetrapods share this strategy with us but overall it is a pretty strange adaptation. Many parts of the environment interact with genes to shape our physiology. These influences include diet and stress. One of the earliest signs for the transition to bipedalism is movement of the foramen magnum. This allows the head to sit in a more upright position. Evidence of this may exist in as early specimens as Toumai which lived 6 to 7 million years ago. Later fossils show more conclusive evidence of bipedalism. Judging from the hips of her species, it seems they had a human-like form of locomotion. Attachments for hip abductor muscles are visible in the pelvic processes in us and earlier bipeds. These muscle attachments allow for our easy gait and keep our opposite foot from dragging on the floor when we take a step. Evidence from Lucy’s spine suggests that she had a spine that curved like ours and allowed her to balance her weight on one leg as she took steps.
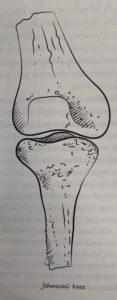
There is some evidence that Lucy had a Johanson’s knee which is a knee that curves inward so that the foot that is on the floor is closer to the center of gravity and allows balance while standing on one foot. A sloping femur and angled knee help to give us the balance we enjoy.
Human ankles are specially shaped to support our weight and are very good at moving along the axis of movement that they need to support efficient locomotion. Fossilized footprints show that hominins were walking upright 3.7 million years ago. Distribution of weight from these footprints corroborates the previous theory that Australopithecus aferensis (Lucy’s species) was able to walk with a fairly efficient gait. We are not as physiologically limited for tree climbing as we may think. If humans climb trees often and from a young age, the soft tissues may form in a way that allows much greater flexibility in the ankle joint and allows us to both walk efficiently and to climb trees similarly to how apes, especially chimpanzees can. We need to be careful to not draw too harsh of conclusions from bones because soft tissues also play a large role in determining functionality of structures.
Roberts cautions against falling into thinking that our mode of locomotion developed linearly through time. She suggests it probably had some other use first then was reapplied to the ground, in contrast to the common narrative of knucklewalking apes beginning to become more and more upright. Roberts suggests that apes that walked in the trees on two legs sometimes were the first step towards upright walking on the ground. Perhaps we are the ones with an old trait and gorillas and chimpanzees have come up with the new mode of transportation in knucklewalking.
Not all of our leg physiology can be explained by walking. In fact, in H. erectus specimens, there is significant evidence suggesting that many of the adaptations favor efficient running. Perhaps this is why our legs are so long, to allow for efficient endurance running. Human adaptations to running are an excellent example of how the environment can shape physiology before genes change. Roberts describes the Baldwin Effect which suggests that variation can arise during the lifetime and these changes can be passed on to offspring in the form of epigenetics. This variation is selected for genetically, but more indirectly, these pressures select for genes that can lead to the desired phenotype. In this way it is important to have genes that are activated to be affected by evolution. Latent potential to be able to pick up new forms of locomotion may be an important factor selected for by evolution even more than direct physiological adaptations.