
If you were to ask yourself the best way to reach the bottom of an Olympic-sized pool, what would first come to your mind? Most of us would probably say swimming upside down or diving on expiration, i.e. exhaling air from our lungs to sink to the bottom.
In this article, we explore the intriguing nature’s response to this question in the specific case of seabirds, capable of diving as well as flying, which inevitably gives rise to biomechanical conflicts between these two types of locomotion. Understanding energy costs in seabird displacement functions is crucial for a deeper grasp of their natural evolution and behavioral habits.
Archimedes, is that you (again)?
Seabirds, when diving, face buoyancy challenges, swimming downwards against the upwards buoyant force. This force is related to the air volume in the seabird’s body and therefore tends to decrease with depth, as the air volume gets compressed due to higher ambient pressures. Over the course of evolution, seabirds have developed unique mechanisms to adapt themselves to the buoyancy change with depth, sometimes at the expense of their flying function.
In the 90s, a study focused on shallow-diving seabirds, specifically ducks, highlighted the existing tradeoff between adaptations for flying and diving. It demonstrated that compared to untrained ducks, dived-trained ducks managed to store significantly more oxygen in their blood and tissues (up to 49.5%) while reducing its presence in the respiratory system (up to 60.7%). This strategic oxygen allocation led to a decrease in air volume, resulting in reduced buoyancy and, subsequently, a lowered diving cost—the energy required for diving.
However, this anti-buoyancy mechanism resulted in greater muscle mass and thus increased body mass, which made flying more energy-demanding and less stable for dived-trained ducks. In other words, it increased their flight cost.
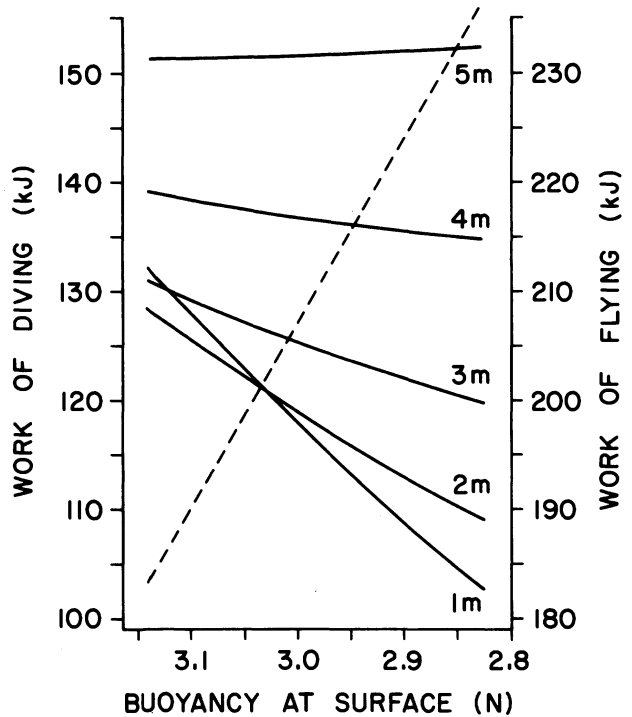
Considering the average daily time spent in the air and underwater and thanks to biomechanical models tuned with experimental data, the researchers were able to produce the figure on the left.
We can observe that the daily work needed for flight (dashed line) rises linearly as duck surface buoyancy decreases. In contrast, daily diving work requirements (solid lines) decrease for depths less than 5 meters. However, beyond 2 meters, reduced diving cost is counterbalanced by increased flight cost, erasing the benefit of the anti-buoyancy mechanism. This confirms the inherent conflict between flying and diving adaptations in seabirds.
Delving into the world of deep-sea divers
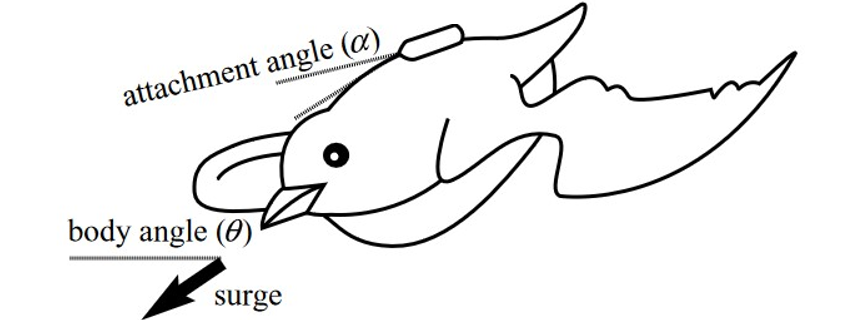
Deep-diving seabirds, whose diving cost isn’t significantly impacted by a strategical repartition of oxygen, developed alternative mechanisms. For example, wing-propelled deep divers adapt their surge frequency with buoyancy, i.e. as the buoyant force decreases with depth, they reduce the frequency at which they produce a force parallel to their body trunk, with the help of their wings.

Among deep divers, the penguin stands out as a subject of great curiosity for researchers. Most penguin species are able to regulate their inhaled air volume depending on the intended dive depth. For example, when they dive very deep, they inhale a large air volume because the buoyancy is negligible where their foraging takes place (deep). Furthermore, it will allow them to ascend partly passively (without using energy), by means of positive buoyancy, which lowers their diving cost.
We tend to forget that penguins are birds, but they are! The optimization of their diving cost through more efficient wing design and on the other hand, the increase throughout time of their flying cost associated with body mass are believed to explain the flightlessness in penguins.
These seabirds may not fly, but their mesmerizing deep-sea dives are a sight to behold: